
鳥学通信 top に戻る


さまよえるコアジサシを守るために必要なこと
文:藤田 剛
自由集会のねらい
「コアジサシは一か所に長居しない...」コアジサシの調査や保全活動をしたことのある人は、そのことを痛切に感じています。景観がそれほど変化していないにも関わらず、ある年には数百羽、数千羽がコロニー繁殖していた場所に、翌年数羽しかやってこないということはよくあることのようです。
これは、コアジサシがある地域に安定して繁殖し続けるためには、何か所もの営巣適地を必要としている可能性を示しています。コアジサシの生息地を保全するためには、どこか一か所だけ営巣地を守るだけでは不充分かもしれないのです。
しかし、このコアジサシコロニ?の不安定さについての定量的な情報はごくわずかしかありません。営巣地を放棄する要因やコロニ?継続期間、その場所を捨てたコアジサシがどの範囲まで移動するのかといったことは、ほとんど報告されていません。
この自由集会では、広域・長期モニタリングデ?タをもとに、コアジサシがさまよう理由や範囲を検討し、コアジサシの営巣地保全のために広域にわたるモニタリングデータが保全にどう重要なのか、3つの講演によって話題を提供し、議論することを目指しました。
講演の概要
演者や企画者以外に、25人の方が参加して下さいました。その「熱い」参加者の中には、各地でのコアジサシ調査や保全活動に携わっていらっしゃる方、NPOやNGOで全国規模の水鳥や鳥類のモニタリング調査を実施・コーディネイトされている方たちも含まれていました。なかなかの顔ぶれです。
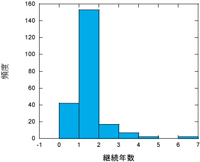
|
| 図. コアジサシコロニーの継続年数 (環境省定点調査のデータをもとに解析、作図) |
1. 日本全域でみたコアジサシの繁殖分布の変遷:藤井幹 (日本鳥類保護連盟)
2. 地域レベルで見たコアジサシの繁殖分布変遷と生息地保全
a. 東京湾:早川雅晴・箕輪義隆 (水鳥研究会)・桑原和之 (千葉県立中央博)
b. 東京湾以外:森田博之・藤田 剛 (東大・農・生物多様性)
藤井氏の講演では、15年間にわたり進められてきた全国的な規模でのモニタリング調査 (環境省定点調査) の結果をもとに、全国的な規模でのコアジサシの現状を解析した結果が示されました。営巣地すべてを完全に把握できていない地域が存在するなど情報に制約があるものの、全国的に個体数が減少している可能性が高いことなどが示されました。

|
| 自由集会参加者 |
森田・藤田の講演では、環境省の定点調査データをもちいた、コアジサシの分散範囲の推定結果が報告されました。まだ試行段階ではあるものの、分散距離は繁殖シーズン内で50km前後、年間で100-200km程度であると推定され、コアジサシの生息地保全にあたって、これらの範囲を考慮することがどう重要なのかを議論しました。
全体議論の概要
これらの講演を受け、まず、コアジサシの広域モニタリングの重要性について議論が始まり、そこから、本種特有のモニタリング実施の難しさなどに話が進みました。営巣地の安定しないコアジサシの調査は、やはり広範囲に行う必要があるにも関わらず、コロニー一か所を見落とすと数100から数1000巣の誤差が容易に生じてしまいます。そのため、非常に大きな労力がかかるという課題が指摘されました。その対策として、精度の粗いデ?タでも解析できる項目を明らかにする必要があること、渡り直前のねぐらなどで個体数を把握することで、繁殖地での調査結果を補正できる可能性があることなどが提言されました。
その後、コアジサシの餌となる魚や捕食者となるカラス類などのモニタリングの必要性と難しさなどについても議論が進み、シギチドリ類や里山鳥類などのモニタリングの話も紹介され、共通の問題点、参考になる点なども議論しました。やがて、コアジサシに限らずさまざまな鳥の広域分布モニタリングの重要性などにも話が及び、熱い議論の中で自由集会が終わり、さらに熱く楽しい2次会への発展したのでした。その2次会の成果は、次回の自由集会に反映されることになりそうです。
謝辞
今回の自由集会の内容の一部は、1988年から環境省が進められてきた定点調査の解析結果に基づいています。この解析にあたっては、環境省および日本鳥類保護連盟にさまざまな便宜をはかっていただきました。

鳥類研究者のための音声分析ガイド
momose
 affrc.go.jp
affrc.go.jpはじめに
パソコンを中心とする情報処理技術やハードウェアの進歩によって、音声、映像、動画などの処理を高速に、かつ手軽に行えるようになってきました。私が初めて購入したパソコン(1982年製のNEC PC-9801)のCPUは5MHzのインテル8086、メモリは640KBでした。私が今使っているDellのマシンは最新型ではありませんが、3.06GHzのCPU2個と4GBのメモリを積んでいます。CPU処理能力はざっと1000倍以上、メモリも5000倍以上に増えています。私は当時のパソコンでプログラムを書いて音声分析を行っていましたが、ウグイスの「ホーホケキョ」という2秒にも満たない鳴き声を分析するのに何時間もかかったものです。何年か後にKAY Sonagraphという専用機を買っていただいたのですが、これは機械式で分析結果を書き込む時にいかにも体に悪そうな臭いのする煙がモクモクと出て、そのたびに同僚から嫌な顔をされたものです。その後KAYにもDSP付きのリアルタイム処理できる機種が出て、とてもうらやましく思いました。現在私の職場には昔何百万円もしたこの機械が置いてありますが、私は触ったことすらありません。今ではパソコンで同じことがずっと容易に行えるので、もはや使う必要がないからです。
さて、前置きが長くなりましたが、この稿では、音声処理について詳しくない方を対象に、パソコンを使った音声分析、特に声紋(Sound Spectrogram)分析の手法について解説します。といっても、ソフトの日本語版マニュアルを書く様なつもりはなく、知っていると役立つ、あるいは知らないと変な間違いをしそうなポイントのみ解説したいと思います。ソフトとしては、コーネル大学が開発した Raven 1.2.1 (Build 27.3 Update 22.3)
http://www.birds.cornell.edu/brp/raven/Raven.html
を例に解説します。これは有料のソフトですが、デモモードで起動すると10分間だけ、無料で動かすことができます。Ravenの前にCanaryというソフトが出ていて、これは現在無料化されていますが、Mac OS 9のみの動作となっています。この環境が使える方は利用されるとよいでしょう。ちなみに、Ravenをダウンロードするとついているマニュアルには、基本から説明したとてもわかりやすい説明が載っています。これも参考にされると良いでしょう。
音声のデジタル化
|
| 図1. 連続的な音圧の変化と離散的なサンプリング |
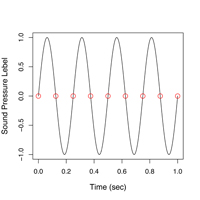
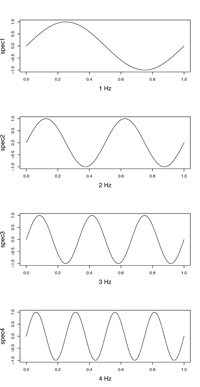
デジタル的に記録できる信号周波数の上限は、サンプル周波数の1/2となります。例えば図2のように、4Hzの音を記録するためには、最低でも8点(8Hz)で記録しないと波の形を捉えることができません。この上限(ここでは4Hz)をナイキスト周波数(Nyquist frequency)といいます。これは理論的な限界値であり、図3のような場合もあり得るので一般にサンプルスピードには余裕を持たせる必要があります。逆に元信号がナイキスト周波数以上の成分を含んでいると、エリアシング(Aliasing)という問題が発生して正しいサンプリングが行えなくなります。
|
|
| 図2. 4Hzの音を記録するには最低8Hzのサンプル周波数が必要 | 図3.とはいえ、こんな場合も… |
音声の一般的なデジタル記録形式である音楽CDのフォーマットは、以下のようなものです。
44.1kHzはおわかりのようにサンプル周波数のことです。1秒間に44.1 x 1000 = 44100回音圧を数値化します。この場合、半分の 22.05kHz までの音声を記録・再生できます。この周波数は一般的な人の可聴域よりかなり高く設定されており、充分な高音質と言えます。16ビット、というのは、デジタル化の深度(Bit depth、サンプルサイズ)で、音圧(電圧)を何段階にわけて記録するかを表しています。この場合2の16乗 = 65536で、±32768 の段階に分けて数値化します。リニア、というのは圧縮をしていない、という程の意味です。パソコンで使われる非圧縮のファイル形式には、ウィンドウズの *.wav、マックの *.aif、ネクストの *.sndなどがあります。これに対し、ポータブル音楽プレーヤーなどで良く使われる *.mp3、AAC(*.m4a)などは圧縮フォーマットです。圧縮フォーマットを絶対使うなとはいいませんが、特に制限となる理由がなければ、オリジナルの音声は上記のCDと同じか、同程度の品質で、wav、aiffなどの非圧縮形式で記録されることをお勧めします。衛星画像の解析をする研究者やプロの写真家が、オリジナル画像をJPEGのような圧縮フォーマットで保存しないのと同じことで、圧縮フォーマットでは信号細部の情報が失われ、それは決して復元できないからです。とはいえ、圧縮が絶対ダメというわけでもありません。もし圧縮をする場合でも、あまり強度の圧縮は避けた方が良いでしょう。
録音機材と、パソコンへの取り込み方
私は鳥学通信第8号で黒田さんが紹介されている(黒田 2006) Roland社製 EDIROL R09というレコーダーを最近は使用しています。これですと、USB接続により簡単にパソコンに音声を取り込むことができ便利です。また、たいていのパソコンにはマイク/ライン入力端子があって、アナログ音声をデジタル形式で取り込む機能がついていますので、アナログで記録された信号でも簡単にデジタルファイルにすることができます。こう書いているだけで腹が立ってくるほど昔の環境とは雲泥の差があります。
声紋 (Sound Spectrogram)とは?
声紋分析について細かく説明する前に、まず声紋とはどんなものかご説明しましょう。まず、次の音声ファイルを再生してみて下さい。
ウグイス (Cettia diphone) 雄のType-Hのさえずりです(Momose 1999)。日本にお住まいの方なら、よくご存じの鳴き声ではないでしょうか。次にどちらかのファイルをダウンロードして、そのファイルをRavenから開いてみて下さい(Windowsの場合はリンクを右クリック、Macの場合はリンクをcontrol+クリックして、リンク先のファイルをダウンロードしてください)。
File / Open Sound Files…
(まず、Fileメニューをクリックして、その下のOpen Sound Files…というメニューを選択し、音声ファイルを探してOKをクリックします。)
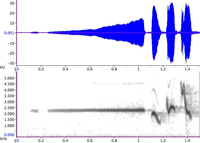
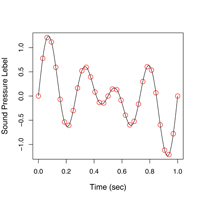
|
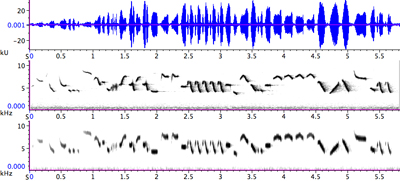
| 図4. ウグイスのさえずり(H型)の波形(上)と声紋(下) |
声紋は、音声信号の周波数分布が時間とともに変化する様子を表すものです。時間は左から右に向かって進行します。周波数は下から上で、高い音程上側に描かれますので、ある意味楽譜と同じように読むことができます。この声紋がどのように描かれるのかを次に解説します。そのためにはまず、第3の表現法である、パワースペクトルについて説明しなくてはなりません。
パワースペクトル(Power Spectrum)について
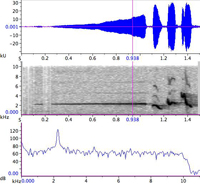
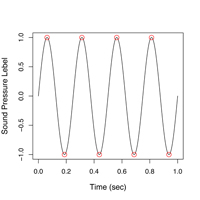
|
| 図5. 0.938秒時点でのさえずりパワースペクトル(一番下の図) |
View / New/ Spectrogram Slice View
を選択すると、図5の様な画面が表示されます。波形表示のグラフのY軸というか、グラフ左端のピンク色の縦線をクリックしたまま,マウスを右に移動させてみて下さい。一番下のSpectrogram Sliceの表示が連続的に変化するのがおわかりかと思います。これが各時点における音声のパワースペクトルで、横軸は周波数、縦軸は音圧(Sound Pressure LevelまたはSPL、単位はdBデシベル)を表しています。周波数軸に沿って、音のエネルギーがどのように分布しているかを示した図です。声紋は、実はこのスペクトルの強弱を周波数軸に沿って濃淡表示した縦に細長い帯を、横(時間軸)方向につなぎ合わせて描いた図に他なりません。要するに、スペクトルが時間に沿って変化する様子を図示している訳です。
パワースペクトルの計算方法
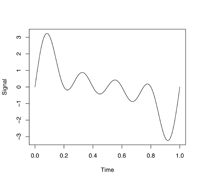
|
| 図6. 複雑な波形も… |
|
| 図7. 単純な波形(正弦波)の和に分解できる |
短時間スペクトルと時間窓
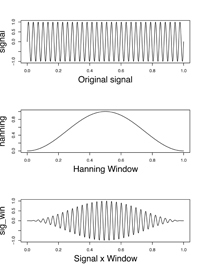
声紋はスペクトルが時間により変化する様子を一目で見られるようにした図だということはおわかりいただけたでしょうか。そこで次に、スペクトルの時間的変化、という問題について考えてみます。ある瞬間における音の周波数というものは、原理的に計測困難です。音とは本来空気などの振動のエネルギーですから、しばらく観測しないと波が上がったり下がったりする様子を見ることはできません。つまり、ある時点の周波数は、その時点の少し前から少し後までの波形から平均的に求めざるを得ません。このため、図8のように、信号を「時間窓」というものを使って少しずつ切り出していき、切り出された短い区間の信号を変換することで、その時点におけるスペクトルを計算します。少し込み入った話になりますが、フーリエ級数は本来無限の長さを持った時間(?∞?+∞)に対して定義されるもので、今話しているような短時間スペクトルを計算する時は、この短時間の波形が永遠に繰り返されるものとして計算します。この際、区間の最後と(次の区間の)最初に不連続があるとノイズが生じてしまうため、図8の真ん中のグラフの様な形をした「窓関数」を掛け合わせて、端を滑らかにしてから分析します。声紋分析の際には、この時間窓の中心点を少しずつずらしながら、スペクトルの変化を計算していきます(これをMoving window法といいます)。Raven では、このあたりの設定を、
View / New / Spectrogram View…
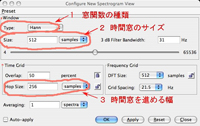
を選ぶと現れる、Configure New Spectrogram Viewウィンドウで設定することができます(図9)。1が窓関数の指定(デフォルトは図8に示したHanning Window)、2が時間窓のサイズで、音声データのポイント数または実際の時間で指定できます。3は、時間窓を1回にどの位前に進めるか、というステップ幅で、デフォルトでは時間窓の半分に設定されています。
周波数分析の設定について
|
| 図8. 窓関数による信号の切り出し |
|
| 図9. 窓関数などの設定法(クリックで拡大表示) |
これは私が以前軽井沢で録音した、ミソサザイ(Troglodytes troglodytes)のさえずりです(Kroodsma & Momose, 1991)。どちらかのファイルをダウンロードしてRavenから開き、Spectrogramを表示してみて下さい。
図10の上(真ん中)は時間窓のサイズを512ポイントに設定した場合、下は64ポイントに設定した場合です。昔は機械的な帯域フィルターを用いて声紋を描いていましたので、その名残で上のような設定をNarrow-band(狭帯域)、下のような設定を Wide-band(広帯域)と呼んでいます。狭帯域では、声紋が周波数軸方向に圧縮されて、周波数分解能が良くなっている代わりに、時間による変化は何となくなだらかに描かれていることがわかると思います。逆に広帯域設定では、時間的変化がシャープにとらえられている代わりに、周波数軸には間延びしていて、周波数分解能が落ちていることがわかります。これは、短時間フーリエ変換を行うことにより生じるトレードオフです。
|
| 図10. 時間窓の長さを変えた分析例。上は512ポイント(Narrow-band setting)、下は64ポイント(Wide-band setting) |
信号の前処理
|
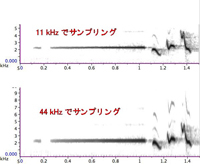
| 図11. サンプル周波数を下げると、周波数分解能が「見かけ上」上がる。 |
ポイントは、「サンプル周波数を下げる」ことです。周波数分解能はナイキスト周波数/時間窓のサイズ(ポイント数)ですから、同じ時間窓サイズで考えた場合、サンプル周波数が半分になると、周波数分解能も倍、というか、より細かい周波数を区別できることになります。図11は、ウグイスのさえずりを11kHz(上)と44 kHz(下)でサンプリングしたものを同じ条件で分析した結果です。上の図の方が、「ホー」の部分の線が細く、周波数分解能が高いことがわかると思います。
最初の方で私は、「なるべくCD音質で録音しておけ」みたいなことを書きました。それと矛盾したことを言っていると思われるかも知れませんが、そうではありません。オリジナルの音声は、記録としての価値もありますので、なるべく情報量が多く、手を加えない形で保存することを勧めます。が、分析の際は、その中から必要充分な情報だけを抽出して行った方が色々都合良いことがあるのです。そのために必要となるのが信号の前処理です。次の様な手順で信号を調整して、分析を行って下さい。
それぞれの処理を行う意味は、ここまで注意深く読んで来られた方ならお分かりになると思います。さて、上記処理の1から4までは Raven で行えます。4を行うには、
Edit / Filter / All with / New Filter…
を選択し、出てきた画面でBandpassをチェック。周波数範囲に、上の例なら
From: 0、To: 22050
と入力します。
上記5については、Raven ではできないようなので、別のソフトを利用して下さい(もしかしたらできるのかも知れませんが)。無料のソフトである、AppleのiTunesを例に方法を説明します(Mac版のiTunes 7.0.2)。
iTunes/環境設定/「詳細」タブ/「読み込み」
を選んで、読み込み方法に例えば「WAVエンコーダ」、設定を「カスタム」とすると、サンプルレート(サンプル周波数)、サンプルサイズ、チャンネル数などが設定できます。その後、変換したいファイルを
ファイル/読み込み
で読み込んだ後、ファイルをマウスで選択してから
詳細/選択項目を WAV に変換
とすると、一括して変換を行ってくれます。この後、再びRavenに戻って、処理6を続けて下さい。
録音の状態や機材によっては、音声に直流(DC)成分や低周波の雑音などが含まれている場合があります。分析対象の音声信号と雑音の帯域が異なる場合は、この前処理の段階でフィルター処理によって除去すると良いでしょう。雑音の帯域が例えば500Hz以下だとすれば、上記の処理4での「From:」の値を 600とかに設定することで除去できます。この適切な値の設定には多少の試行錯誤が必要です。この場合の注意点は、フィルター処理は、やるならなるべく一回、あるいは必要最小限の回数で済ませる、という点です。うまく行かないと思ったらフィルターをかけ直すのではなく、必ず前の状態に復帰してからやり直して下さい。あまり何回もフィルターをかけていると、分析対象の信号構造にも悪影響が出てきます。
声紋の計測方法について
最後に、声紋分析の結果から計測を行う際の注意点について述べます。声紋から時間、周波数などを読み取って計測したい、という場合があると思います。Ravenには、このための便利な機能が色々付いています。一つ守っていただきたいことは、「周波数の計測はパワースペクトル上で行う」という点です。理由は図5を見ていただければわかると思います。スペクトルでは、周波数のピークがピンポイントで読み取れますが、声紋は、このレベルを任意の段階でスライスして濃淡表示しただけのものですから、レベルの設定で声紋パターンの巾も変わるため、正確な計測は難しくなります。
終わりに
他にいくつか解説したいこともあったのですが、取りあえず声紋分析の方法については、一通り概略的な解説ができたと思います。参考にしていただき、色々な分野で研究を進めて下さることを期待しています。
引用文献ほか
・黒田治男 (2006) 鳥類の音声研究のためのデジタル録音機材. 鳥学通信 No.8
http://ornithology.jp/osj/japanese/katsudo/Letter/no8/OL8.html#03
・Kroodsma, D.E. & Momose, H. (1991) Songs of the Japanese Population of the Winter Wren (Troglodytes troglodytes). Condor 93:424-432.
以下のリンクからPDFをダウンロード可能
http://elibrary.unm.edu/sora/Condor/issues/v093n02/index.php
・Momose, H. (1999) Structure of Territorial Songs in the Japanese Bush Warbler (Cettia diphone). Mem. Fac. Sci., Kyoto Univ. (Ser. Biol.) 16(2): 55-65.
・作図に使用したRのコード (R_Scripts_Momose.txt)
Further Reading
・Hopp, SL, Owren, MJ & Evans, CS (Eds.) (1998) Animal Acoustic Communication. Springer, Berlin. ISBN 3-540-53353-2
・Kroodsma, DE & Miller, EH (Eds.) (1996) Ecology and Evolution of Acoustic Communication in Birds. Comstock / Cornell, Ithaca and London. ISBN 0-8014-8221-6

タイの調査事情
はじめに

|
| 図1. オオサイチョウのオス(2002年2月7日撮影) |
カオヤイ国立公園
カオヤイ国立公園はタイで最初の国立公園として1962年に設立されました。タイの東北部に位置し(101°05’-50E, 14°05’-15N)、標高250?1,351m、面積は2,168 km2のタイで5番目に大きな保護区です。年平均降水量は2,360mm (1993-2002)で、大きく4月?10月の雨季と11月?3月までの乾季にわけられます。

|
| 図2. キタカササギサイチョウのオス(2001年12月3日撮影) |
カオヤイで現在進行中の研究
カオヤイ国立公園では1970年代後半からバンコクにあるMahidol University所属のPilai Poonswad教授が率いるサイチョウチーム(Poonswad et al. 2005)や同じくマヒドン大学のWarren Brockelman教授が率いるテナガザルチーム(Brockelman et al. 1998)による長期生態研究が継続されている場所です。テナガザルチームの調査地であるMo Sing Toプロットには30ヘクタール(500 - 600m)の長期生態研究サイトが設定されています。このサイトでは現在King Mongkut’s UniversityのGorge Gale博士を中心として、小型鳥類の繁殖生態の研究が進められており(Nimnuan et al. 2004, Pierce et al. 2004)、現在はノドジロカンムリヒヨドリ(Alophoixus pallidus)の研究が集中的に行われています。
調査許可の申請手続き
外国人研究者がタイ国内で調査を行う場合には、National Research Council of Thailand(NRCT)からの許可を得ることが必要です。調査地が保護区(国立公園や野生生物保護区など)である場合、その管轄を行っているNational Park, Wildlife and Plant Conservation Departmentにも別の申請書を提出する必要があります。これらの調査許可手続きに関する情報は全てNRCTのホームページから情報を得ることができますので、タイでの調査を考えている方はまずこちらにアクセスしてみてください。申請手続きがスムーズに行われた場合でも、許可がおりるまでに3ヶ月程度はかかるのが普通ですので、余裕を持って許可申請を行う必要があります。めでたくNRCTからの調査許可を取得することができたら、日本国内のタイ大使館で3ヶ月間有効のシングルエントリーの調査ビザを取得することができます。以前は日本国内で一年間有効のマルチエントリーの調査ビザを取得することができたのですが、現在は3ヶ月を超えて長期滞在したい場合は、バンコクの入国管理局でビザの延長申請をする必要があります。
参考URL
National Research Council of ThailandのInformation for Foreign Researchers
http://www.nrct.net/eng/index.php
National Park, Wildlife and Plant Conservation Department
http://www.dnp.go.th/National_park.asp
タイ王国大阪総領事館(東京総領事館のサイトには調査ビザのページがない)
http://www.thai-kansai.net/jp/consulate-service/visa/consular-japanese-6.html
タイでのバードウオッチング情報
http://www.thaibirding.com/index.htm
参考文献
・Brockelman WY, Reichard U, Treesucon U, Raemaekers JJ (1998) Dispersal, pair formation and social structure in gibbons (Hylobates lar). Behav Ecol Sociobiol 42:329-339.
・Lynam AJ, Round PD, Brockelman WY (2006) Status of birds and large mammals in Thailand's Dong Phayayen - Khao Yai forest complex. Biodiversity Research and Training (BRT) Program and Wildlife Conservation Society, Bangkok, Thailand.
・Nimnuan S, Round PD, Gale GA (2004) Structure and composition of mixed-species insectivorous bird flocks in Khao Yai National Park. Nat Hist Bull Siam Soc 52:1-9.
・Pierce AJ, Pobprasert K, Gale GA (2004) Breeding ecology and nesting success of Abbott's Babbler (Malacocincla abbotti). Wilson Bull 116:275-277.
・Poonswad P, Sukkasem C, Phataramata S, Hayeemuida S, Plongmai K, Chuailua P, Thiensongrusame P, Jirawatkavi N (2005) Comparison of cavity modification and community involvement as strategies for hornbill conservation in Thailand. Biol Conserv 122:385-393.

鳥類調査研究所ヘルゴランド分室(Institut fur Vogelforschung, Vogelwarte Helgoland)

|
| 写真1. 鳥類調査研究所(IfV)ヘルゴランド分室の全景. |

|
| 写真2. ヘルゴランド本島北西端にあるThe Long Annaと呼ばれる岩. |

|
| 写真3. 崖で繁殖するシロカツオドリとミツユビカモメ.ウミガラス類は7月上旬に巣立つので、この時期にはいない. |
|
| 写真4. 捕獲施設(ヘリゴランド・トラップ)の模式図.追い込み用の小径が点線で示されている. |

|

|

|
| 写真5. ボランティアの勢子の後を追いかける. | 写真6. 捕獲網室内の小鳥を捕まえる.網室のドアを抑えているのがシュラム技官. | 写真7. 捕獲箱の説明をするヘルゴランド分室長のヒュポップ博士. |

|
| 写真8. 計測小屋の様子.ボランティアが手際よく、足輪をつけて、計測し窓から放す。 |

|
| 写真9. カラーリングで標識されたクロウタドリ幼鳥.この個体には、発信器が付いている. |
ヘルゴランドは、日本の旅行ガイドにはまったく紹介されていませんが、ドイツ、デンマーク、オランダから観光客がたくさんやってくる海岸リゾートで免税店がたくさんあります。本土から船が着く11時をすぎると島の人口は2,3倍に膨れあがり観光客であふれかえります。船が出港する3時半から人が減り始め、4時半にはほとんどの店が閉まり人通りが少なくなります。海岸には、ゼニガタアザラシがごろごろ寝転がって、港ではケワタガモが潜水して蟹を食べていました。IfVの研究に興味がある方ばかりでなく、バードウォッチャーも渡りの時期に訪れれば、探鳥+リゾート気分を味わえる一押しの穴場だと思います。ハンブルグに行く機会がありましたら、お出かけください。
・Huppop & Huppop (2006) Aussagekraft der Helgolander Fangzahlen uber grosraumige Bestandstrends. Jber. Institut Vogelforschung 7:8.
・Hill & Huppop (2006) Techniken zur Erfassung des ?unsichtbaren Vogelzugs“ uber See. Jber. Institut Vogelforschung 7:21-22.
・Sacher et al. (2006) Brutvorkommen und Zugverhalten der Amsel auf Helgoland. Jber. Institut Vogelforschung 7:11.

2007年最初の鳥学通信をお届けします。今号の「鳥類学の最前線」には、百瀬さんによる、音声分析の大変詳しい解説記事が掲載されています。このような技術解説は、なかなか学術雑誌には掲載されません。しかし、この手の情報は、プロ・アマ問わず、研究者にとって大変有用なものです。「野外調査のTips」と合わせて、このコンテンツは鳥学通信のウリのひとつと考えています。「○○手法やテーマついて(○○さんに)解説して欲しい」、というリクエストがありましたら、是非編集部までお寄せください(副編集長)
鳥学通信は、皆様からの原稿投稿・企画をお待ちしております。鳥学会への意見、調査のおもしろグッズ、研究アイデア等、読みたい連載ネタ、なんでもよろしいですので会員のみなさまの原稿・意見をお待ちしています。原稿・意見の投稿は、編集長の永田宛 (mailto: ornith_letterslagopus.com) までメールでお願いします。
鳥学通信は、2月,5月,8月,11月の1日に定期号を発行します。臨時号は、原稿が集まり次第、随時、発行します。
編集・電子出版:日本鳥学会広報委員会