2016年8月9日
独立行政法人 国立科学博物館
植物研究部多様性解析・保全グループ
杉田典正
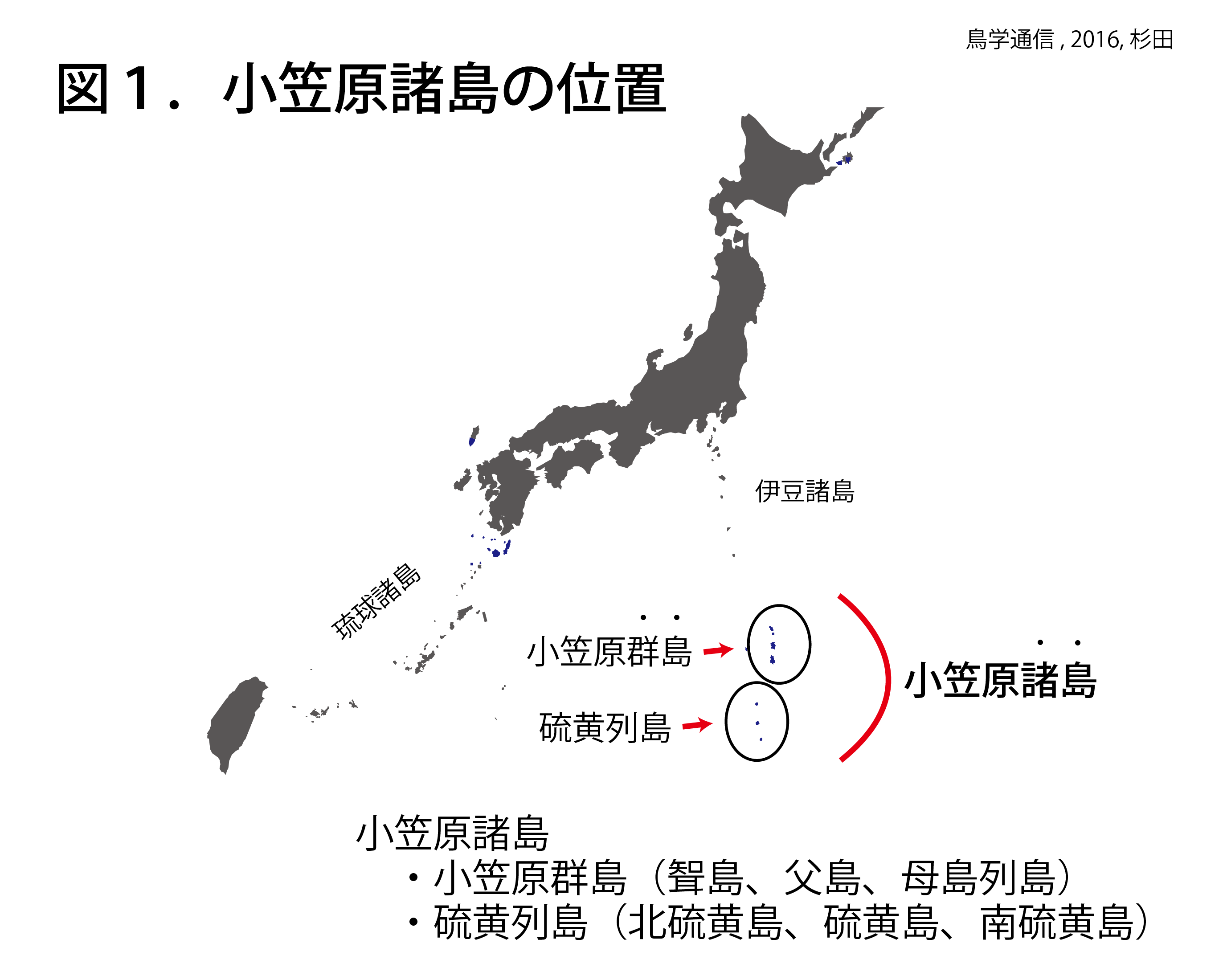
小笠原諸島は、東京から南に約1000 kmから1200 kmの南の海上に浮かぶ小さな島々の集まりである(図1)。小笠原諸島は、父島、母島などを有する小笠原群島と北硫黄島、硫黄島、南硫黄島の3島から成る硫黄列島の2つの群島から構成される。小笠原諸島へは、およそ週に一度東京から出港するおがさわら丸に乗って24時間かけて行く方法しかない。母島へ行くには、父島で船を乗り換えてさらに2時間半かかる。もっと南の硫黄列島へは旅行で行くことはできない。小笠原諸島は日本で一番遠い場所のひとつかもしれない。
小笠原諸島は、海洋島である。海洋島は、島が出来てからほかの地域と一度も繋がったことがない島のことをいう。海洋島は、独自の生態系がつくられる。小笠原諸島も、他の地域では見られない独特の海洋島生態系ができている。例えば、オガサワラオオコウモリは小笠原諸島唯一の固有哺乳類で、小笠原で独特な行動と生態を進化させてきた。小笠原は、独自の生態系と進化を観察できることが評価され、2011年には世界自然遺産に登録された。
小笠原群島と硫黄列島は、どちらも海底火山に由来する。しかし、島の形成時期が全く異なる。小笠原群島は、約4500万年前の海底火山を起源にする古い島である。一方、硫黄列島は、数万年内に噴火した海底火山と隆起により形成された新しい島である。小笠原群島は、伊豆諸島やマリアナ諸島など周辺の島嶼地域の中でもっとも古い。
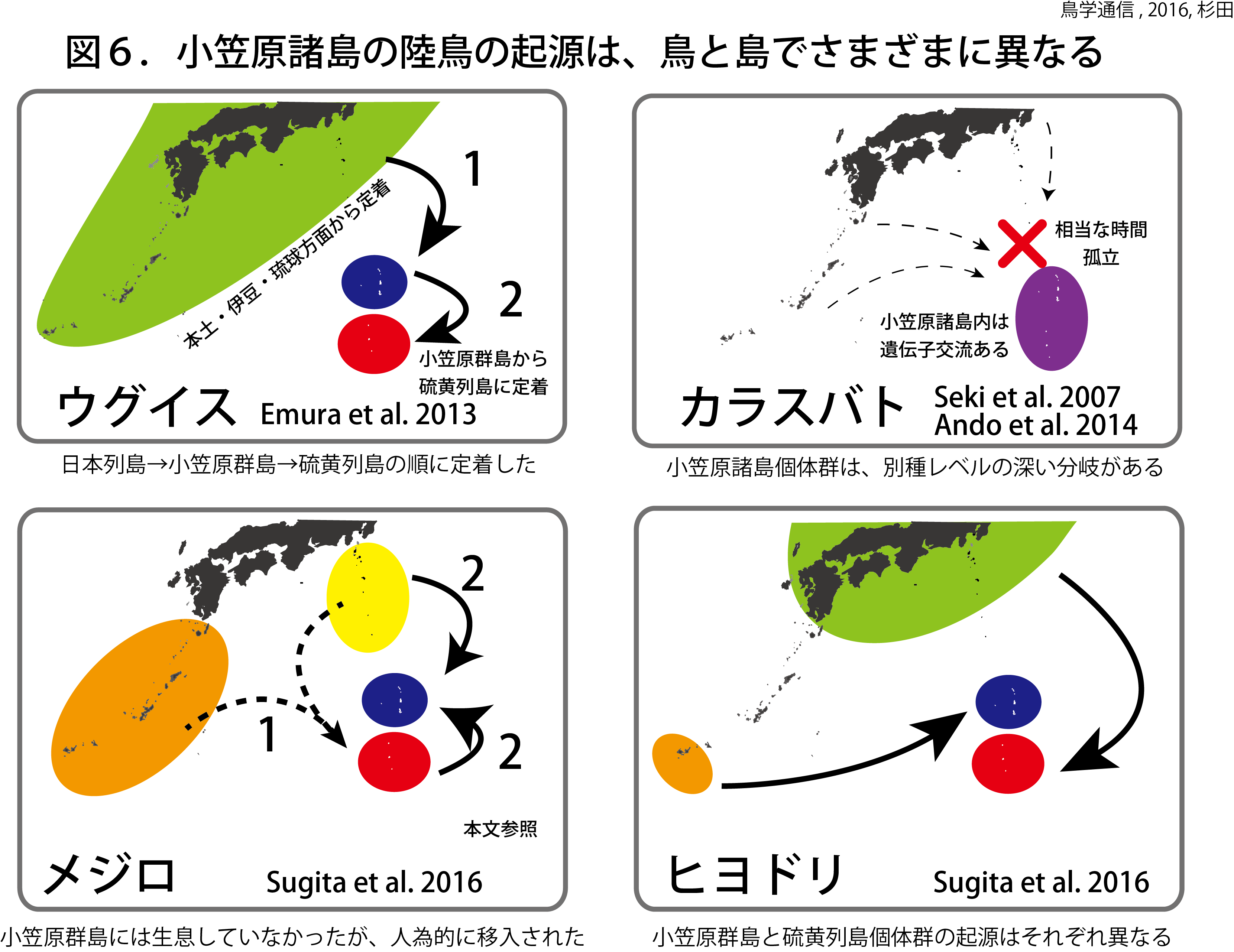
小笠原諸島で繁殖する陸鳥の数は少ない。10種の陸鳥が繁殖するのみである。過去にはオガサワラマシコやオガサワラガビチョウ、オガサワラカラスバトなど小笠原諸島固有の陸鳥が生息していたが、絶滅した。メグロは、母島列島で生き残っているが、父島列島と聟島列島で絶滅した。現在、小笠原群島と硫黄列島の両群島に共通して繁殖する陸鳥は、カラスバト、ヒヨドリ、ウグイス、カワラヒワ、トラツグミ、イソヒヨドリの7種類に過ぎない。カラスバトとカワラヒワは、本州の個体群と大きな遺伝的差異があり、ミトコンドリアDNAで別種レベルの深い分岐がある。ヒヨドリとウグイスは、日本列島の個体群のミトコンドリアDNAと比べて数塩置換の違いである。これら5種の小笠原諸島個体群は、亜種に分類されている。トラツグミは、1930年に初めて小笠原諸島で発見され、1960年代頃から増え始めた。ミトコンドリアDNAのCOI領域では、遺伝的差異は見つからなかった。モズは父島で1980年代から繁殖していたが、2000年代の中頃に絶滅したとみられる。
小笠原諸島の陸鳥は、大陸から遠く離れた島であるため個体群間の交流が少なく遺伝的に分岐が進んだ。一方、面積の小さい海洋島では、環境が不安定であり、人口学的要因などによって定着しにくいという特徴もある。小笠原諸島の陸鳥の鳥類相の変遷は、海洋島の特徴をよく表している。
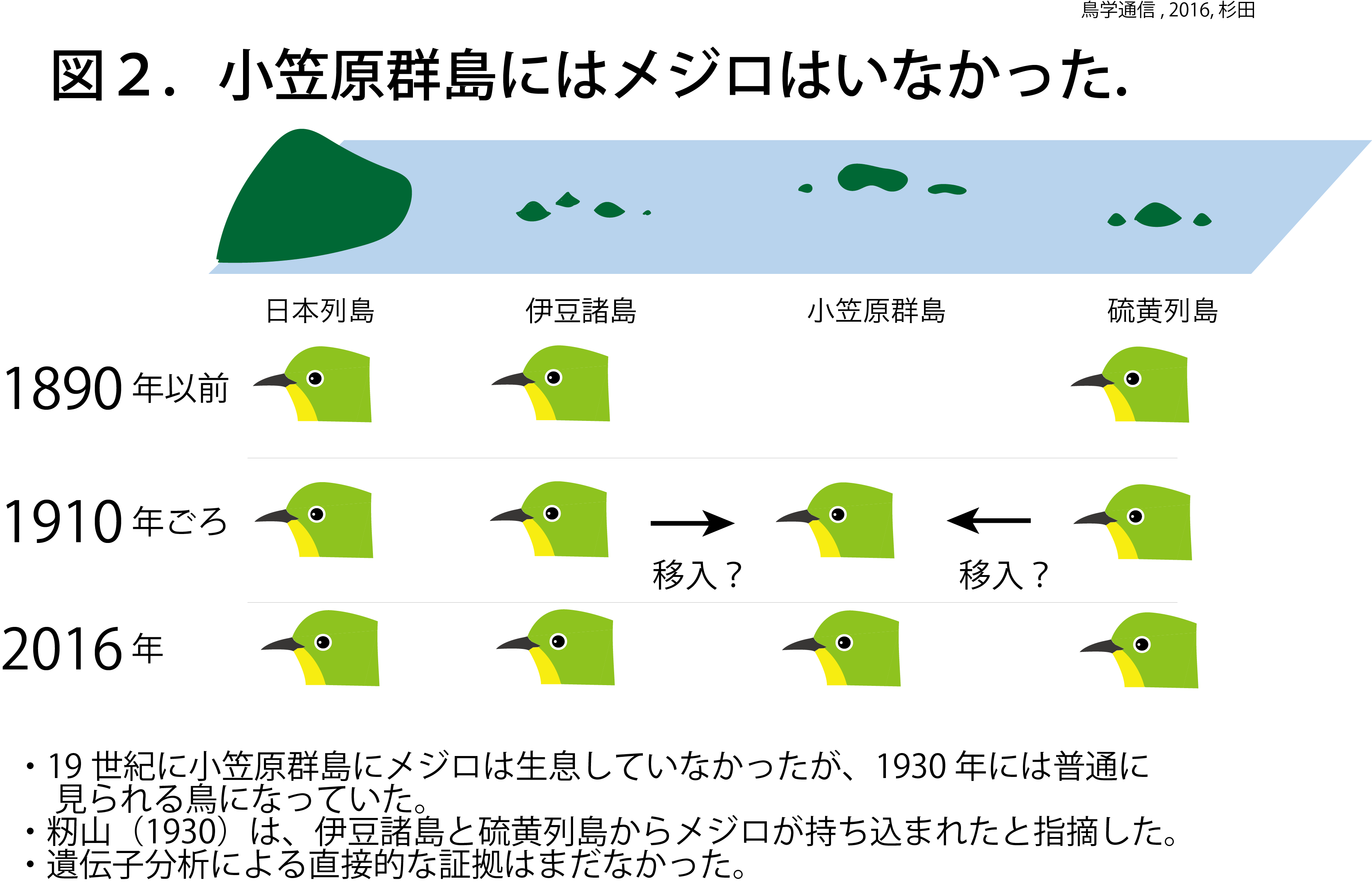
問題のメジロである。日本鳥類目録第7版(2012)によると、小笠原諸島のメジロの分布は、硫黄列島のみresident breederとなっている。父島や母島に行ったことのある方は、「おや?」と思うかも知れない。小笠原諸島の父島や母島でメジロをたくさん見たはずだと。確かにメジロは、現在の小笠原群島にたくさん生息している。もっとも目に付きやすい鳥だ。しかし、今から約百数年前、1900年以前には小笠原群島にはメジロは生息しなかった(図2)。過去の記録でも明らかである。1827年イギリス軍艦ブロッサム号で父島に上陸したBeechey艦長は、ヒトを恐れない黄色いカナリアの様な鳥(おそらくメグロ)を観察したが、メジロは記録していない。1890年の研究でも、小笠原群島でメジロは採集されなかった。ところが1930年になると、メジロは父島で最もよく見られる鳥になっていた。籾山徳太郎は、1930年に発表した論文で、島民に聞き取り調査を行い、1900–1910年頃に硫黄列島と伊豆諸島からメジロが持ち込まれたと考えた。同時に、彼は、小笠原群島のメジロの形態形質に亜種イオウトウメジロとシチトウメジロの中間的特徴があることから、小笠原群島個体群は両亜種の交雑個体群であると示唆した。日本鳥類目録第7版では、小笠原群島のメジロはintroduced breederとされている。しかし、小笠原群島のメジロが本当に硫黄列島と伊豆諸島から持ち込まれたのか、DNA分析による証拠はまだ無かった。
DNA分析に使える血液サンプルが硫黄列島のメジロで採られていなかった。硫黄列島は、北硫黄島と南硫黄島は無人島、硫黄島は自衛隊基地のために上陸調査が難しい。特に、南硫黄島は、原生自然環境保全地域に指定されていること、島の全周が切り立った崖で出来ており上陸しにくいことから、学術調査隊が長い間立ち寄っていなかった。だが、一連の硫黄列島の学術調査に参加してきた川上和人博士(森林総合研究所)によって、硫黄3島すべてからメジロ個体群の血液サンプルが得られた。小笠原諸島のメジロの遺伝分析と他の地域との比較が可能になった。
この記事では、小笠原諸島のメジロのミトコンドリアDNAを分析してその起源を調べた研究を紹介する。この記事は、Zoological Scienceで発表された小笠原諸島のメジロとヒヨドリのミトコンドリアDNA分析を行った研究内容にもとづく(Sugita et al. [2016] Zoological Science 33: 146–153)。小笠原諸島のヒヨドリのDNA分析の解説については、BIRDER 2016年7月号(文一総合出版)と森林総研プレスリリース資料をご覧頂きたい。本研究のすべての分子実験は、国立科学博物館の実験室で行った。
独自に進化した硫黄列島のメジロ
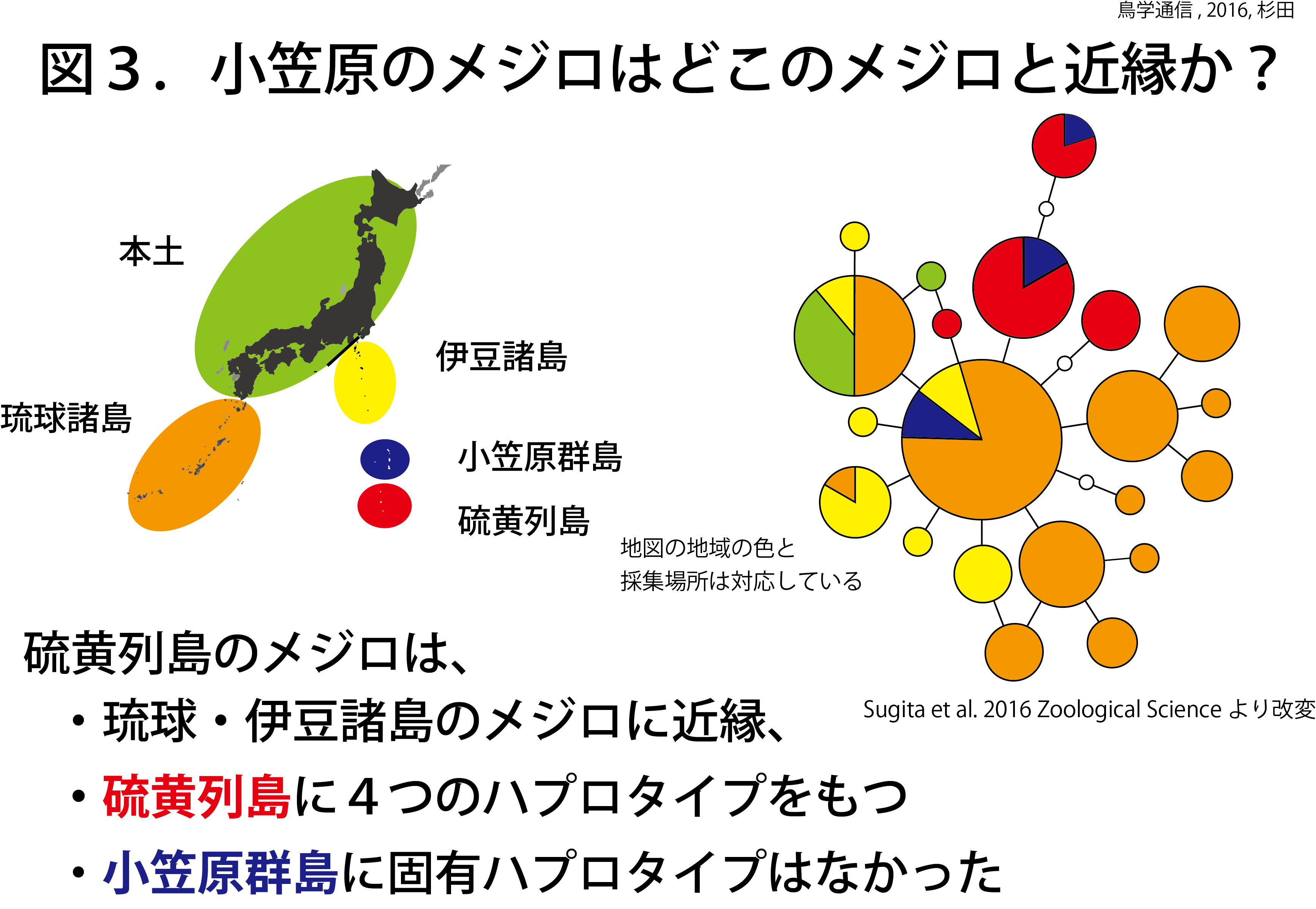
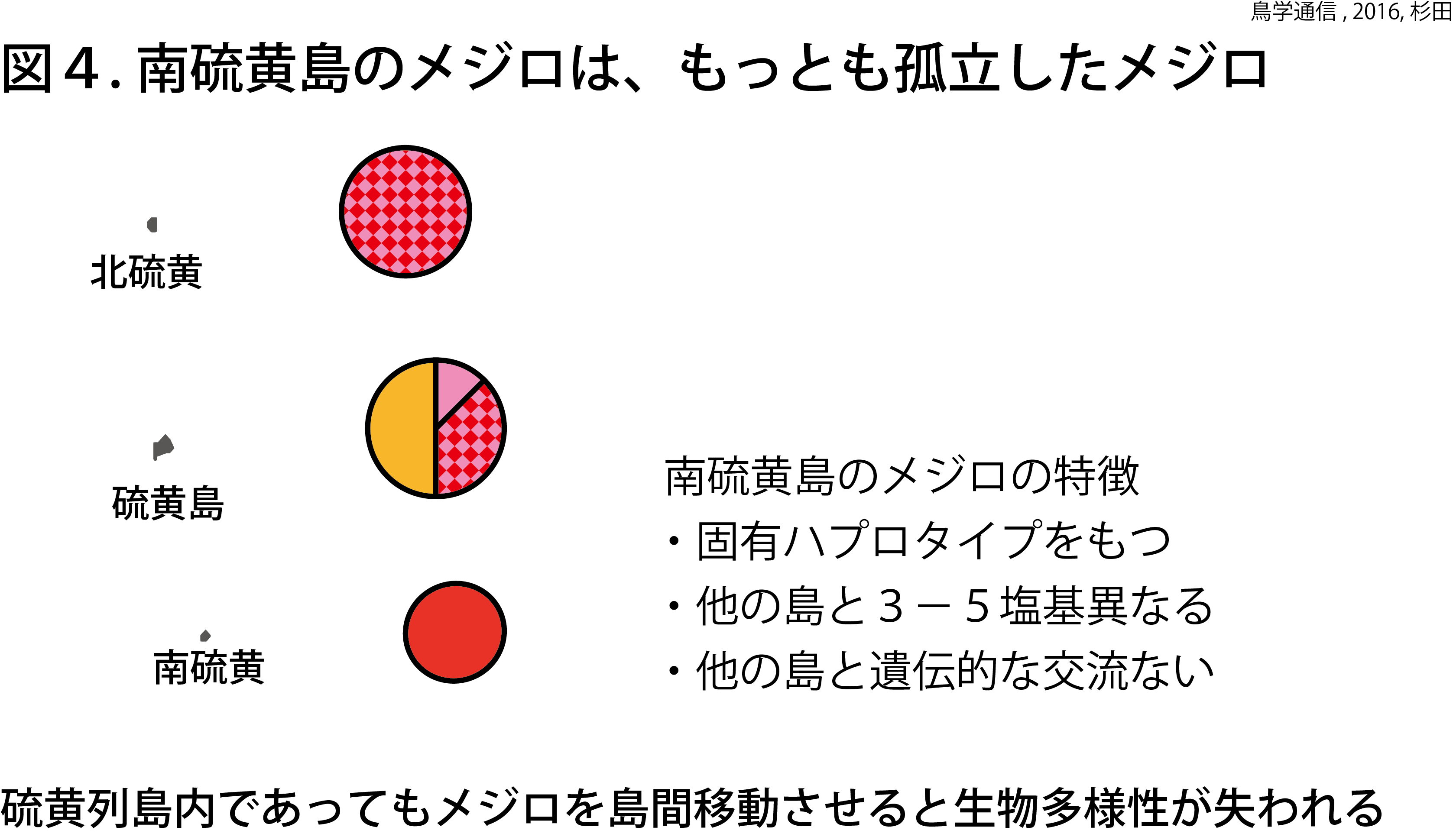
硫黄列島のメジロの解析のために、ミトコンドリアDNAのチトクロームオキシターゼcサブユニット1(COI)遺伝子のDNA配列を使用した。ミトコンドリアDNA多型をハプロタイプという(異なるハプロタイプ同士は配列が異なるということ)。図3に赤色で示されている部分が硫黄列島のメジロ個体群のハプロタイプで、4種類見つかった。硫黄列島のメジロ個体群のハプロタイプは、小笠原群島のメジロ個体群を除いて(後述)、他の地域から見つからなかった。硫黄列島の3島を詳しく見ると、南硫黄島の個体群のハプロタイプは他のどの島からも見つからない南硫黄島固有のハプロタイプであった(図4)。
これらの結果は、硫黄列島のメジロ個体群は、伊豆諸島や琉球諸島のメジロと近縁の関係であることを示している。硫黄列島のメジロは、独自のハプロタイプをもった遺伝的に分化した個体群であることも示している。南硫黄島の個体群は、他のどの島とも遺伝的な交流のないもっとも孤立した個体群であることがわかった。硫黄列島のメジロは、祖先集団が硫黄列島に定着した後、それぞれの島で移動性を低下させて、交流のない独立した集団になったと考えられる。生物進化の見本として、小笠原諸島の世界自然遺産としての価値をさらに高める研究成果となった。
生物多様性は、外来種と在来種との交配などによって容易に低下する。硫黄列島で独自に進化したメジロ個体群は、小笠原諸島の海洋島生態系における取り替えの効かない重要な要素である。特に南硫黄島のメジロ個体群は、他のどの島とも遺伝的に異なる。硫黄列島のメジロの保全は、島単位で考えるべきだ。
連れてこられた小笠原群島のメジロ
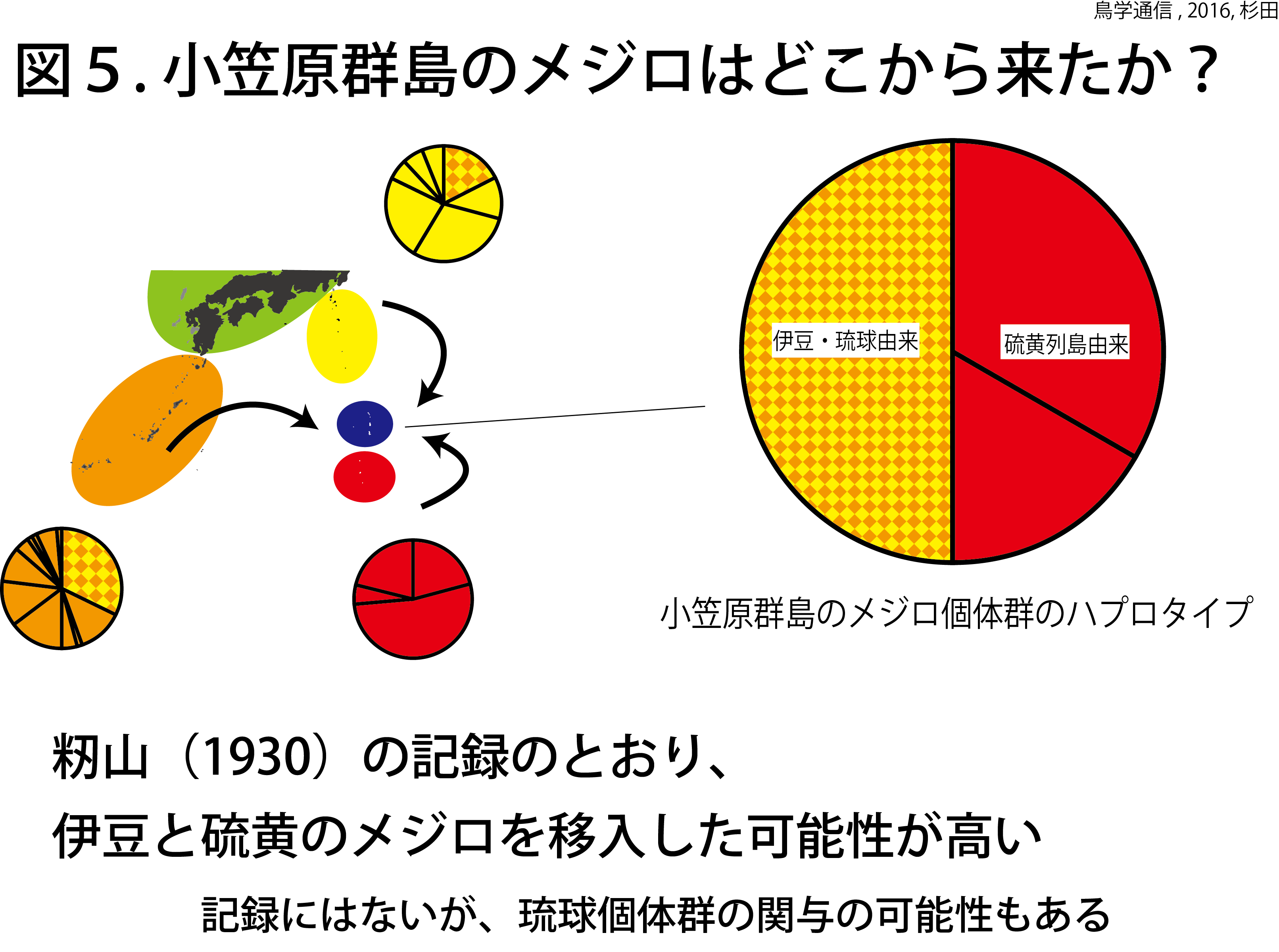
小笠原群島のメジロ個体群に固有のハプロタイプは無かった(図3)。小笠原群島のメジロ個体群から3つのハプロタイプが発見されたが、これらは琉球諸島と伊豆諸島、硫黄列島の個体群と共通のハプロタイプであった(図5)。硫黄諸島では、固有ハプロタイプが見つかったことと対照的である。この結果は、メジロが小笠原群島に自然分布していたのではなく、近年外部からメジロが小笠原群島に移入されたことを示唆している。自然分布していたのなら、小笠原群島にも硫黄列島のメジロのように固有ハプロタイプが発見されるはずだ。この見方は、過去の記録とも一致する。
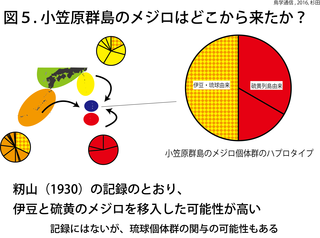
伊豆諸島のメジロ個体群は琉球諸島個体群の一部と同じハプロタイプを共有していたものの、籾山の聞き取り調査の結果などを考慮すると、籾山が1930年に発表した仮説の通り、小笠原群島のメジロ個体群は硫黄列島と伊豆諸島から来たと考えるのが妥当だろう。ただし琉球諸島のメジロの関与も否定できない。今回、籾山の仮説以来86年間謎であった、小笠原群島のメジロの起源を現代のDNA分析技術により検証することができた。籾山は、小笠原群島のメジロがシチトウメジロとイオウトウメジロの交雑個体群であるという仮説も提唱した。これについては、今後、核DNAによる詳細な研究が必要である。
様々なルートでやって来た小笠原の陸鳥
小笠原諸島の陸鳥の起源や島間の遺伝的関係性は種ごとに異なる。いくつかの種で小笠原群島と硫黄列島間の遺伝的関係が知れられている。ヒヨドリは、小笠原群島と硫黄列島の個体群でそれぞれ異なる起源をもつ(図6)。ウグイスは、琉球と伊豆諸島以北の個体群から小笠原群島に進出した。その個体群の一部が、さらに南の硫黄列島に進出したようだ。アカガシラカラスバトは、カラスバトと別種レベルの遺伝的差異がある。しかし、小笠原諸島内では、アカガシラカラスバトは、小笠原群島と硫黄列島を行き来している。小笠原群島のヒヨドリは八重山諸島を起源にする。一方、硫黄列島のヒヨドリは本州などの個体群を起源にする。それぞれ時間的にずれて小笠原諸島に定着した。その後互いに交流しなかったようだ。これらの研究結果は、島の形成時期、定着時期、生態的特性、島の群集構造などの違いにより、島によって異なる生物相が成立することを示している。
小笠原諸島の世界自然遺産への登録以来、自然再生事業が行われている。小笠原諸島では、鳥のような飛べる動物であっても、島や列島ごとに鳥類相や遺伝構造が異なることがわかった。この結果から、島や列島を単位とした保全計画が必要と言える。
参考文献
Ando H, Ogawa H, Kaneko S, Takano H, Seki S, Suzuki H, Horikoshi K, Isagi Y (2014) Ibis 156: 153–164.
Beechey F (1832) Narrative of a Voyage to the Pacific and Beering’s Strait, to Co-operate with the Polar Expenditions: Performed in His Majesty's Ship Blossom, under the Command of Captain F. W. Beechey, R. N. in the Years 1825, 26, 27, 28. Carey & Lea, Philadelphia, USA.
Emura N, Ando H, Kawakami K, Isagi Y (2013) Pacific Science 67: 187–196.
籾山徳太郎 (1930) 日本生物地理学会会報 1: 89–186.
日本鳥学会(2012)日本鳥類目録第7版, 日本鳥学会, 三田.
Seebohm H (1890) Ibis 32: 95–108.
Seki SI, Takano H, Kawakami K, Kotaka N, Endo A, Takehara K (2007) Conservation Genetics 8: 1109–1121.
森林総合研究所(2016)絶海の孤島、小笠原の鳥はどこから来たのか?2016. Apr. 7. URL: www.ffpri.affrc.go.jp/press/2016/20160407/
杉田典正(2016)BIRDER 30(7): 34–35.
Sugita N, Kawakami K, Nishiumi I (2016) Zoological Science 33: 146–153.