天野達也(ケンブリッジ大学)
先月、Successful conservation of global waterbird populations depends on effective governance 「世界の水鳥保全の成否は各国のガバナンス有効性に依存する」と題した論文を発表しましたので、その内容と研究の経緯をここで紹介させていただきます。
論文の閲覧はこちらで、また日本語での解説はこちらもご覧ください。
「生物の数がどこでどのように変化しているのか」という問いは、研究を始めた学生の頃から一貫して興味の中心であったように思います。
私の生物保全に関する研究は、北海道の宮島沼でマガンの数を数えることから始まりました。その後研究を進めるにつれて、ヨーロッパではモニタリング調査で得られたデータの解析によって、様々な鳥類について詳細な個体数の変化が明らかにされていることを知りました。
このように生物の減少を明らかにすることは、科学者が生物多様性保全のために提供できる最も基礎的で根本的な知見のひとつと言えます。私も自然とそういった研究を志すようになりましたが、ヨーロッパ、特にイギリスで蓄積されたデータや知見は非常に豊富で、また当時は2010年目標に向けて世界の生物多様性変化に関する論文が盛んに発表されていたこともあり、「世界のどこでどのくらい生物が減少しているか」という問いは、既に解決済みのようにも感じられました。

宮島沼に渡来するマガン
そのような認識を改めるようになったのは、世界の脊椎動物の個体数変化を示す指標であるLiving Planet Index(LPI)について、あるひとつの図(こちらのFig 5)を見たのがきっかけでした。この世界的な取組みでも、使われているデータは欧米のものに大きく偏っていて、他の多くの地域では未だ生物の数の変化は明らかにされていないということを、この時に強く実感しました。
そこでまず、日本の鳥類モニタリング調査から得られているデータを使って、鳥類の分布や数の変化を明らかにする研究を始めました。そしてこれらの研究を進めていく過程で、鳥類の中でも特に水鳥についてはInternational Waterbird Census(IWC)という枠組みのもと、世界規模で個体数の調査が長年継続されていることを知ったのです。とは言え、自分がこのデータに取り組むようになるとは、当時すぐには想像できなかったのですが、ちょうどこの頃イギリスで一年の在外研究を行っていたことで、共同研究者との議論や周囲からの刺激もあり、これまで自分が行ったことのない大きなことに取り組んでみたいという意欲も高まっていました。そうして、このIWCデータの解析に取組むプロジェクトを計画したのが2010年です。
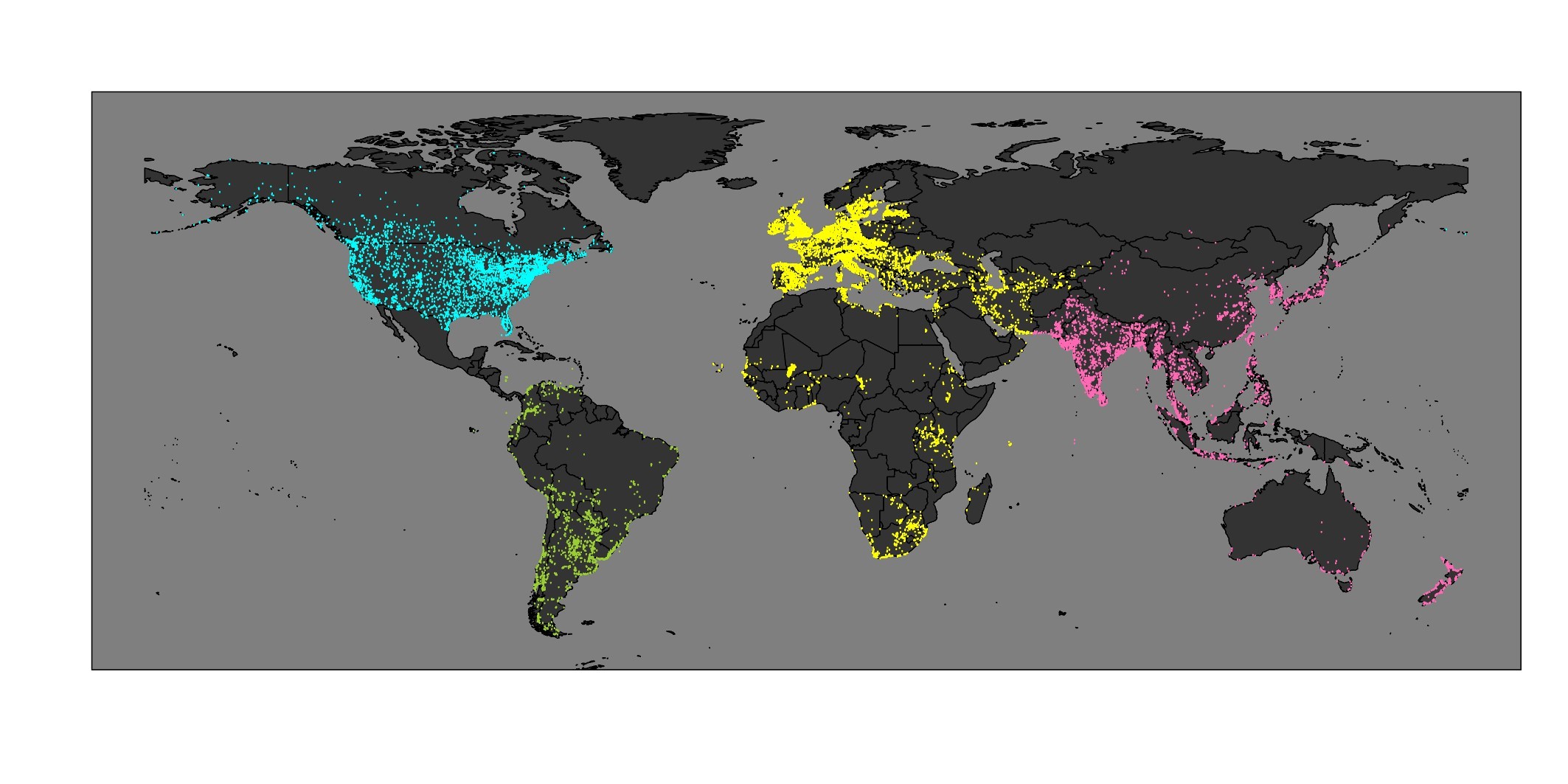
Wetlands International(WI)が行っているIWCは、1967年にヨーロッパで始まり、今では世界180か国、5万地点にも及ぶ調査地で、毎年1月に水鳥の個体数をカウントする、まさしく「世界規模」の調査です。アジア、中東、南米など、先述したLPIでも十分にカバーされていない地域に多くの調査地が存在するのは驚異的で、このデータを用いれば「世界のどこでどのくらい生物が減少しているか」という、長年抱えていたシンプルな疑問への答えに、少しでも近づけるのでは、と直感したことをよく覚えています。
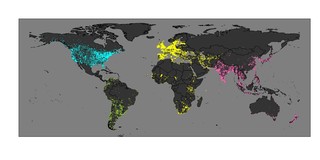
IWCによる調査地(黄色:African-Eurasian Waterbird Census, ピンク:Asian Waterbird Census, 黄緑:Neotropical Waterbird Census, カリブ諸島で行われているCaribbean Waterbird Censusはここでは図示されていない)とChristmas Bird Countによる北米の調査地(水色)

サウジアラビア(左)とオマーン(右)での水鳥調査の例(写真:Szabolcs Nagy)
共同研究者の紹介で、同年に行われたワークショップでWIとの共同研究の話を進め、翌2011年には同プロジェクトをテーマに掲げたJSPS海外特別研究員制度で本格的に渡英…と、ここまではとんとん拍子でした。しかしその後、何年にも渡って幾多の壁にプロジェクトの進行が阻まれることになります。
まず苦労したのは、世界中のデータを自分の手元に集めることでした。IWCはWIによって管理されてはいるものの、人員・資金不足などのため全てのデータが本部に集約化された形にはなっていませんでした。IWCを各地域で構成しているAsian Waterbird Census(アジア)、Neotropical Waterbird Census(南米)、Caribbean Waterbird Census(カリブ諸島)それぞれの担当者とデータ利用を交渉し、必要に応じて各国の責任者にも許可を取ってもらうというプロセスには膨大な時間がかかりました。さらにIWCではカバーされていない北米で利用できるデータを調べ、毎年IWCと同時期に行われているChristmas Bird Countのデータを用いるために、全米オーデュボン協会と共同研究を確立しました。
次に直面した壁は、手に入れたデータの質を管理する作業です。世界各国で集められた500以上もの種のデータを、一種ずつ既知の分布情報と照らし合わして明らかなエラーを排除し、また、国や団体によって異なる種名表記や亜種の扱いを統一していくといった過程は、地道で且つ時間のかかる作業でした。世界で絶滅に最も近いとされているある種が、とある国で数百羽も記録されていたのを見たときには、絶望的な気持ちになったものです…。
またそうして集めて生データには、年によって調査が行われていなかったり大きな測定誤差が含まれていたりという、長期モニタリングデータが抱える典型的な問題が含まれていました。そのため生データを眺めているだけではなかなか個体数の変化を捉えることはできません。
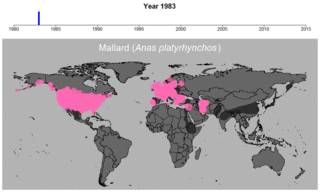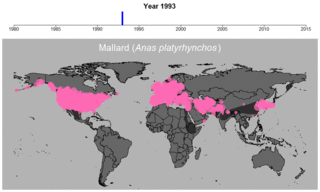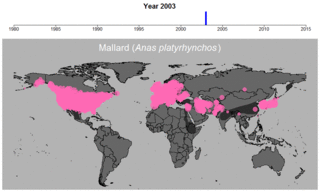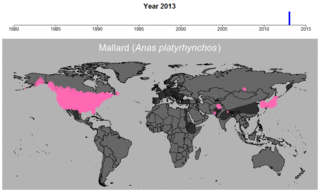
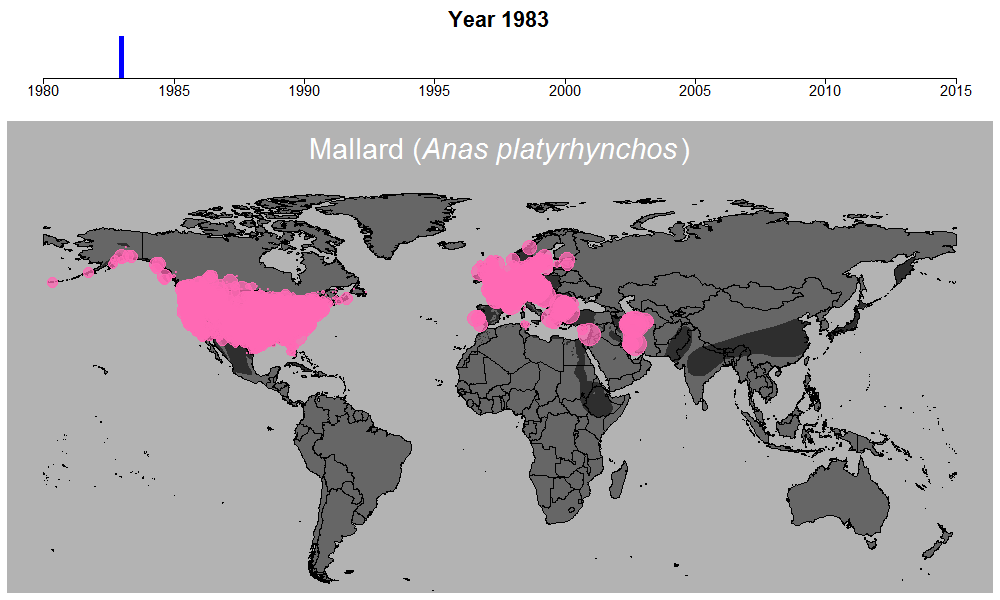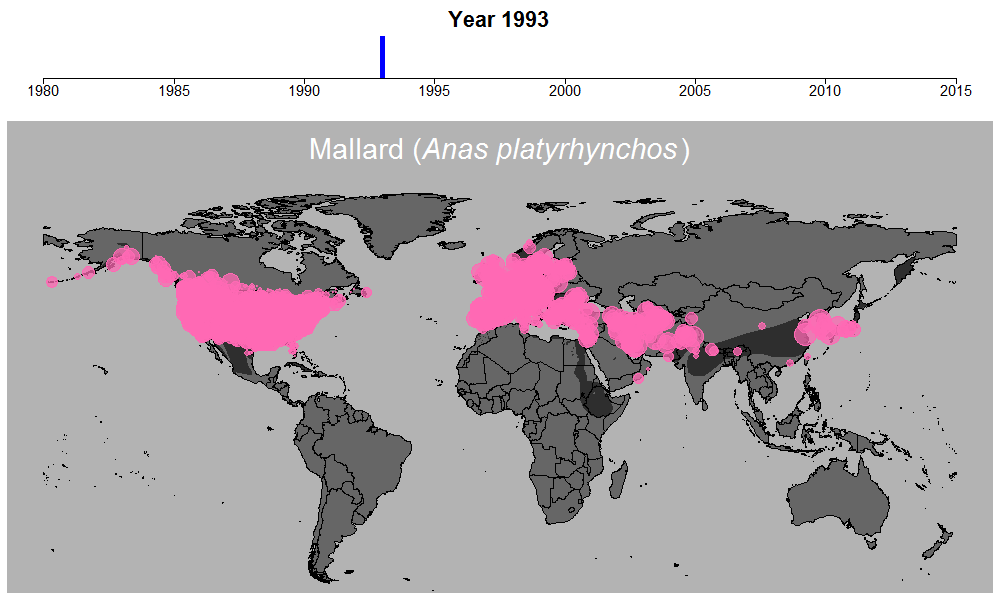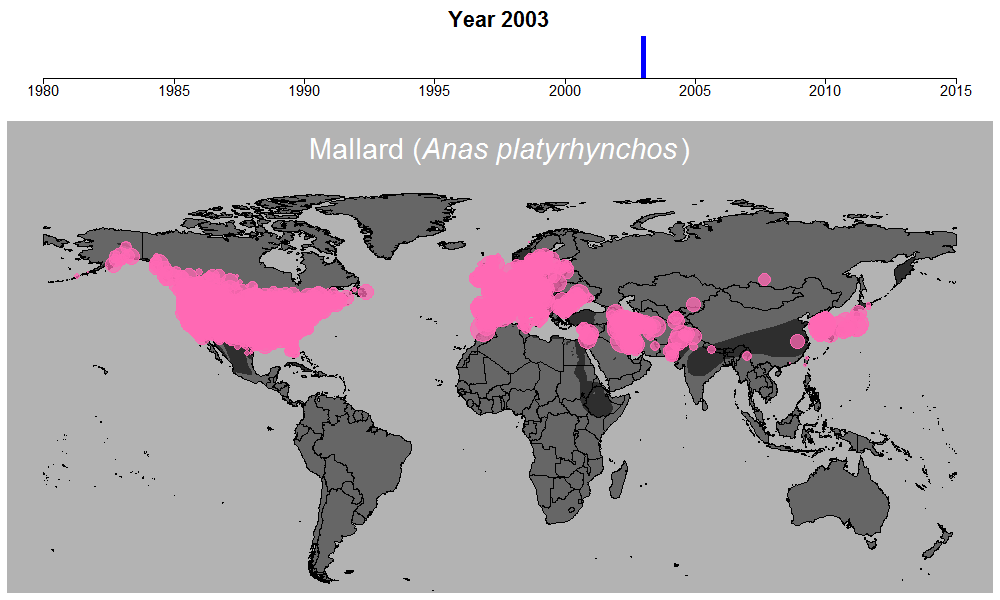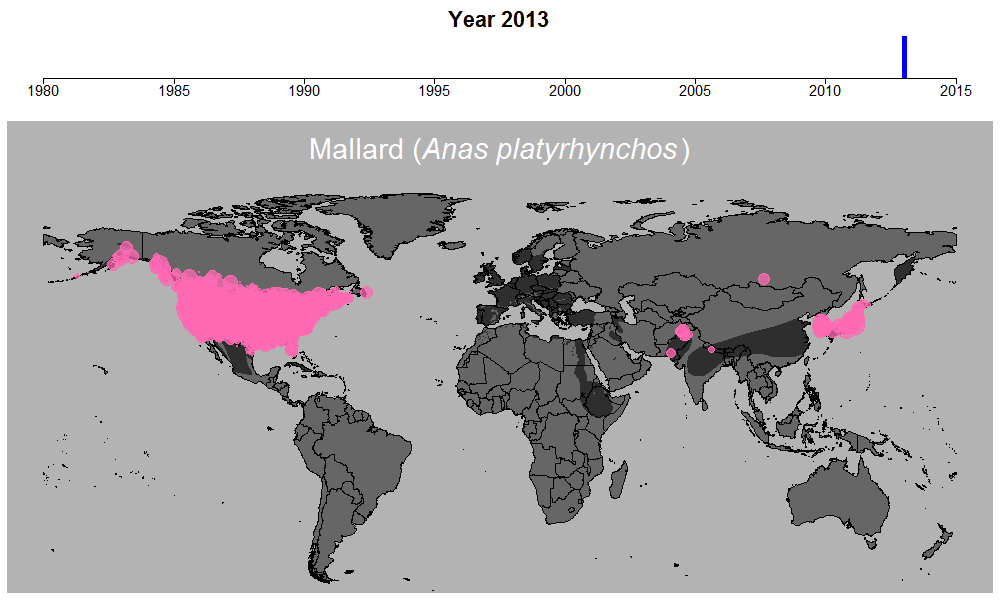
マガモの生データの例。円のサイズは観察された個体数を表す。
これらの問題を考慮するために適切なモデリングが必須でしたが、このモデル計算を行う過程も非常に時間のかかるものでした。この時ばかりは世界中膨大な数の調査地で収集されている水鳥データの存在に感謝しながらも、同時に若干恨めしくも思いました。何せ最もデータ数の多いマガモ一種だけでも、世界1万以上の調査地で12万件以上ものデータが集められているのです。マガモ一種のモデル計算を終えるのには2週間近くかかり、全体の計算時間は数か月以上にも及びました。しかしこういった作業の結果、ついに461種について、世界のどこで、どのくらい数が減っているのか、また増えているのか、明らかにすることができたのです。
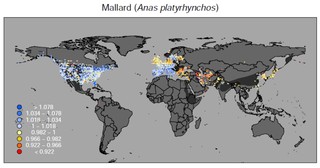
マガモの例。その他の種はこちらで公開しています:https://doi.org/10.6084/m9.figshare.5669827.v1
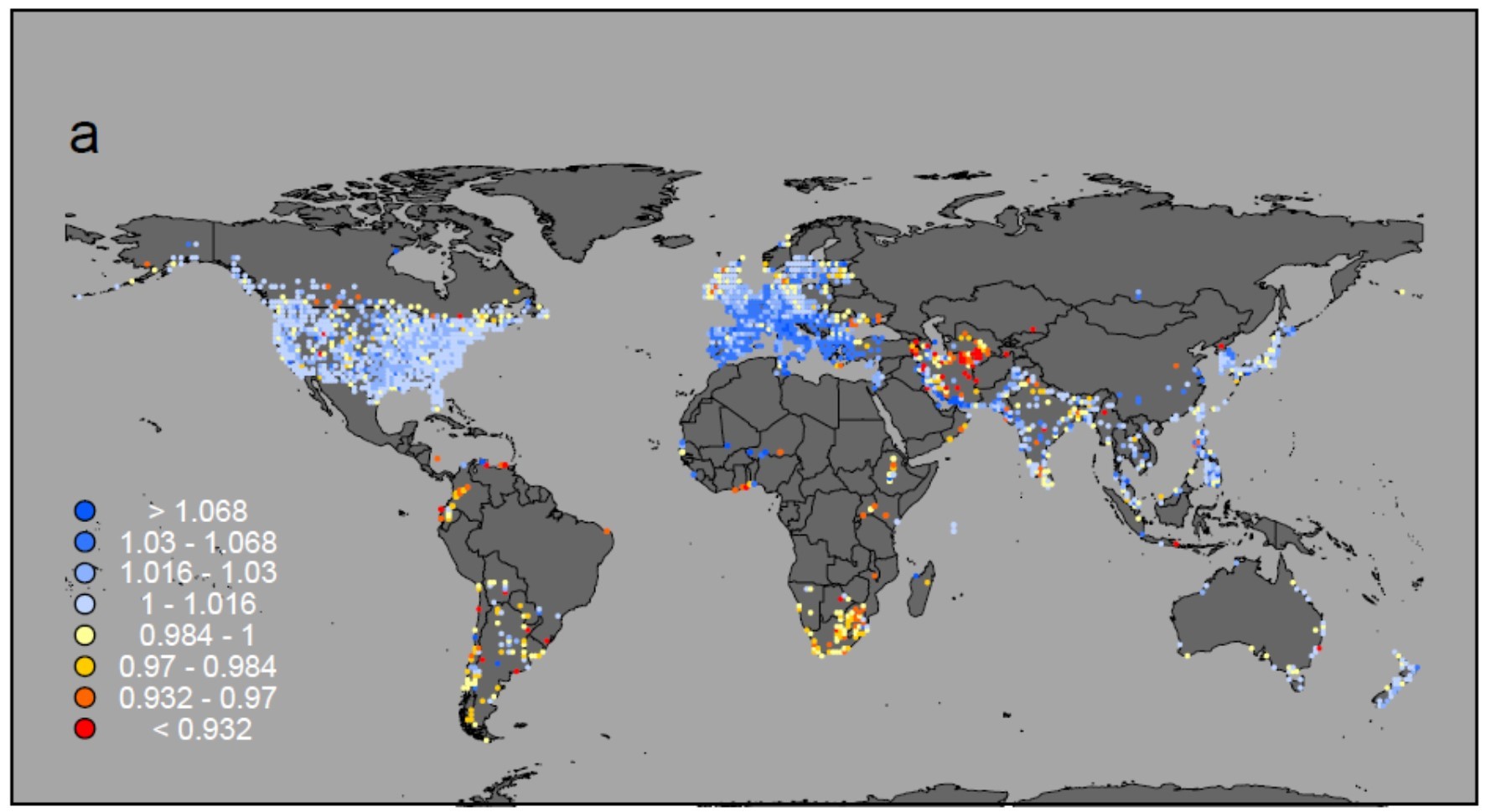
この時、プロジェクトの着想から既に5年近くが経過していました。2015年、当時のオフィスで初めて全461種の個体数変化の地図を重ね合わせ、水鳥全体での変化を表す地図を作製した日のことは、今でも鮮明に覚えています。渡り鳥の減少がよく知られているオセアニアや、生物多様性全般のホットスポットである熱帯地域で水鳥の減少が著しいと考えていたものの、予想に反して最も深刻な減少が見られたのは、イランを中心とした西・中央アジアでした。
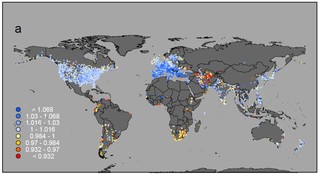
解析に用いた全461種の平均個体数変化
次の自然なステップとして、明らかになった水鳥の増減を何が説明するのか探索しました。国間での生物多様性損失の違いを説明する要因としては、経済発展のレベルや保全に費やした予算などが挙げられていますが、サハラ以南アフリカよりも西・中央アジアで減少が著しいなど、経済レベルのみでこのパターンが説明できないことは明らかです。一方、同時期に関わった学生のプロジェクトで、各国のガバナンス(法律の施行などを通してどれだけ効果的に各国が支配されているか)が生物多様性保全に関わる様々なパターンを説明することが明らかになってきていました。そこで、湿地環境の変化、農地の拡大、気候変動などの人為的な脅威、保護区の存在やガバナンスの程度といった保全の効果に関わる要因、そして渡りの有無や分布域の広さ、体重など各種の特性、という3種類に区分される要因の影響を検証することにしたのです。
解析の結果、ガバナンスの重要性は明らかでした。水鳥群集全体で見た場合、最も減少が著しかったのは経済レベルが低い国ではなく、ガバナンスの有効性が低い国でした。また種間の傾向を見ると、保護区によって保全されている種ほど増加していましたが、この傾向はガバナンスが効果的な国(ヨーロッパ諸国など)のみで見られたのです。一方、ガバナンスの有効性が低い国では、保護区による保全は水鳥の増加にはつながっていませんでした。これらの結果は、ガバナンスという社会政治的な要因が、今や世界全体の生物多様性変化のパターンを作り出すほどに大きな影響力を持っていること、そして、保護区が本来の目的を果たすためには、ただ設置されるだけではなく適切に管理される必要がある、ということを示しています。
これらの結果を様々な人と議論した結果、特にイランを中心とした西・中央アジアでの水鳥に関する情報を、多く手に入れることができました。この地域では歴史的に水鳥の資源利用が行われてきましたが、近年伝統的な猟法に大規模な狩猟がとってかわり、局所的な推定でも毎年数十万羽の狩猟圧が、保護区の内外や種の保全状態を問わずあることが報告されています。

イラン、Fereydunkenarにおける大規模なかすみ網猟 (写真:Petri Lampila)
また過度の水資源利用による湿地環境の消失も知られており(保護区に設定されているイランのLake Urmia のうち、京都府全域に相当する面積が干上がった例)、保護区が設置されていてもこれら複数の直接的な脅威が水鳥の著しい減少につながっていることが推定されます。
ガバナンスが世界的な水鳥の個体数変化を説明するというのは私自身にとっても意外な結果で、当初はなかなか確信を持てないでいました。しかし、上述したような結果をサポートする情報や、2003年にアフリカでの生物多様性とガバナンスの関係を発表し、同じ研究室に所属しているBalmford教授からも心強い意見をもらい、自信をもって論文を書き上げることができました。
その後、完成した論文の投稿直前に共同研究者からデータに含まれたエラーを知らされ打ちひしがれたものの、何とか気持ちを奮い立たせ、さらに再解析に数か月を費やしたのが一年前の年末年始。半年ほどの査読・改訂の期間を経て、ついに着想から7年が経過した昨年末、この論文を発表するに至りました。
この論文は幸運にもNature誌で発表することができました。10年前には自分の論文がNature誌に掲載されるとは考えもしていなかったので、純粋に嬉しく思っています。一方で同じ10年の間に、これら著名な雑誌で発表される論文は、世の中に存在する多くの重要論文の中で氷山の一角のような存在であることも実感してきました。数多くの論文の中から「海上に露出」するためには、運のような自分では制御できない要因も一定の役割を果たすでしょう。また、「海面下」(もちろん論文は世に出ている時点で全て「海上」なのですが)には、同様に重要で新規性の高い論文も数多く存在しています。自分の周囲の人たちがこういった雑誌に論文を投稿している過程を見て、また自分でも何度かその過程を実際に経験することで、海面というただ一つの境界線がその後の見栄えを左右する問題、またそういった論文に付随する著者の様々な思いも、身をもって実感してきました。共同研究者のSutherland教授からは、以前から「(これらの雑誌も)所詮話題性が欲しい雑誌の一つに過ぎないから」と言われていたのですが、今後少しでもそういった境地に達し、氷山全体を見渡す目をもって研究を続けていきたいと思っています。
無論、今回の論文の発表がこの研究の終わりではありません。IWCデータの利点を活かした研究は、共同研究者も含めて今後も推進していくつもりです。また幸いなことに、共同研究を行ったWIはラムサール条約などの関連会議や、各国の調査コーディネーターとも強いコネクションを持っています。この研究の成果を土台とした各地での保全活動の普及や政策への提案、また更なるモニタリング体制の確立など、次の動きは既に始まっています。先述したイランのLake Urmiaでは、日本政府からのサポートも含めた国際的な保全活動の結果、近年その水位レベルは回復傾向にあります。この事例は、暗鬱とした話題が多い環境ニュースのなかで、国際的な保全活動が成功している好例として、もっと注目を集めるべきでしょう。今後、環境変化の影響だけでなく、こういった保全活動の効果を評価する際にも、全世界の水鳥モニタリング調査は重要な役割を果たしていくはずです。

トルクメニスタンの研究者と本研究の結果について議論するSzabolcs Nagy氏。
2010年にWIのワークショップに参加して、ヨーロッパにおける生物多様性変化を評価する取り組みに改めて感銘を受け、「次の10年で、日本を初めとしたアジアでもこういった取組みを進めていくために少しでも貢献していければ」と感じました。その成果を出すのにこの10年の大半を費やしてしまいましたが、今回の成果を弾みとして、8年前に立てた目標をさらにつき進めていきたいと思います。