喧嘩のあとにはフォローが大事 ~セキセイインコのなかなおり~
京都大学 物質―統合システム拠点(iCeMS)
科学コミュニケーショングループ
特定研究員 一方井祐子
突然ですが、友人と喧嘩してしまったらどうしますか?ごめんなさいと頭を下げたり、握手をしたり。ハグをしたりする人もいるかもしれません。では、喧嘩に負けてしょんぼりしている友人を見かけたら?肩をたたいて「元気出せ!」といったり、黙ってご飯に誘う人もいたりするかもしれません。
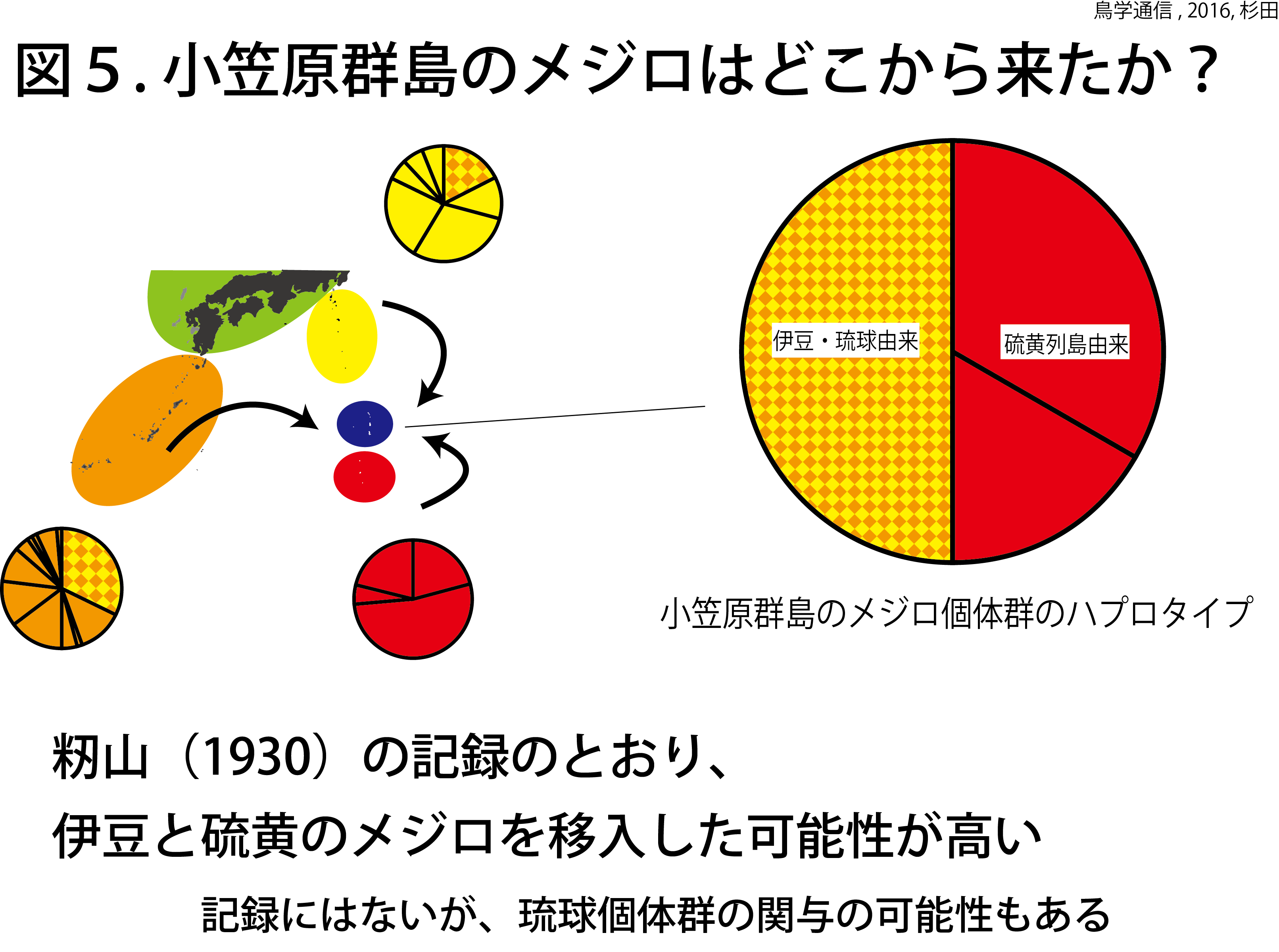
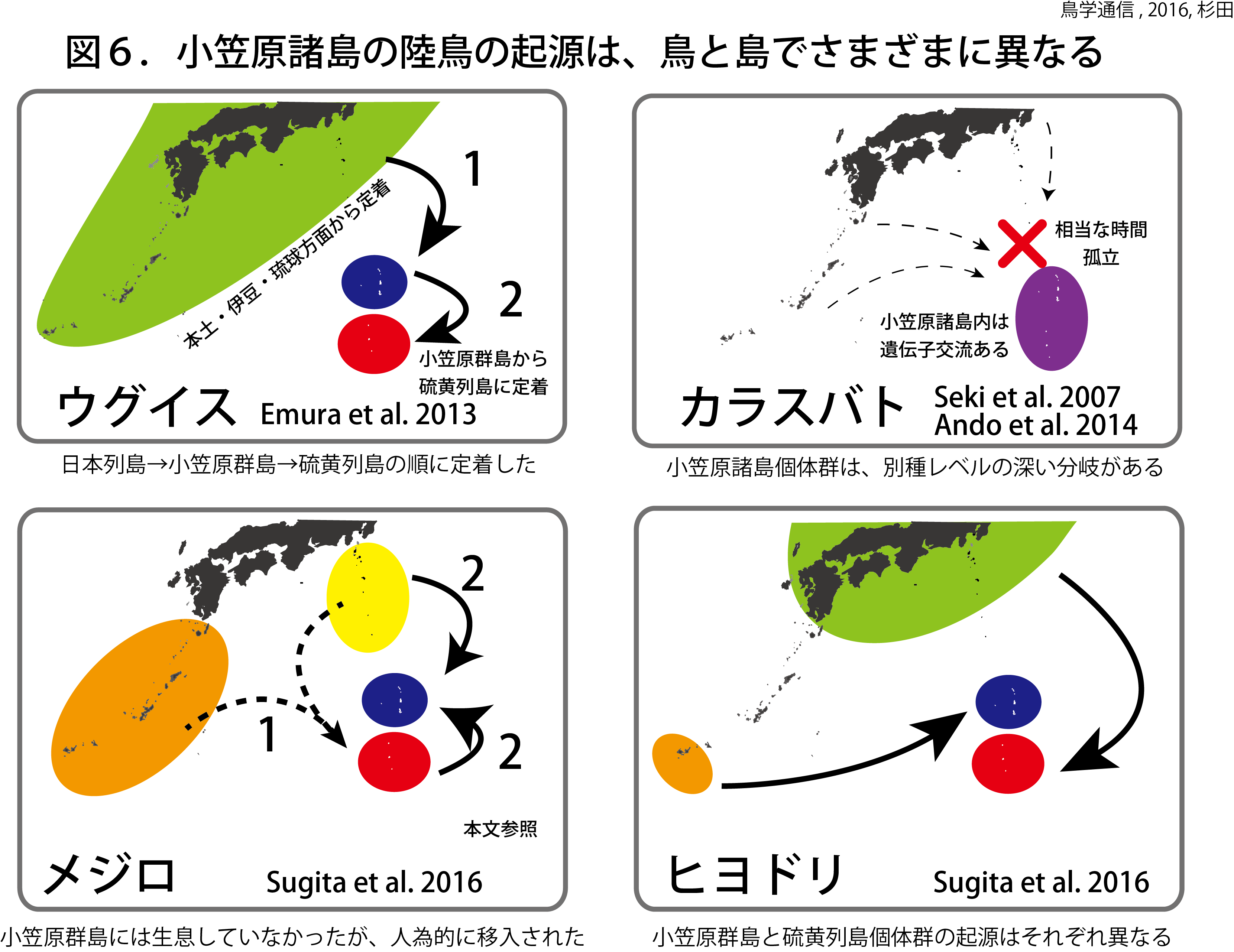
前者の行動は「なかなおり」、後者の行動は「なぐさめ」と呼ばれる行動です。私たちの研究 [1] によって、じつはセキセイインコもこれらの行動をするらしいということが分かってきました。
セキセイインコはおもしろい
私はセキセイインコ(図1)の視聴覚コミュニケーションを研究してきました。ペットとして広く知られているセキセイインコですが、ヒトによく似ているところがたくさんあることはご存じでしょうか。一夫一妻を長く続けるところ。オスとメスの両方が子育てに関わるところ。鳥類にはめずらしく、オスよりもメスの方が攻撃的(!)というところも面白いところです。

図1 セキセイインコ
セキセイインコ同士であくびが伝染すること [2]、セキセイインコの発声パターン(=コンタクトコール)が群れやつがいで似てくること(e.g. [3];[4])も報告されています。これらの行動もヒトとよく似ています。
セキセイインコの喧嘩とは?
今回、私たちが注目したのはセキセイインコの「喧嘩」と「喧嘩のあとに起こること」です。当時、私が所属していた研究室ではセキセイインコの集団飼育をしていました。セキセイインコは多くの時間をつがい相手の近くで過ごします。一方で、なぜだかよく喧嘩もしていました。
嘴を大きく開いて相手を威嚇したり、大きな声をあげながら相手を嘴でつついたり。つがい相手と喧嘩したり、つがい相手以外と喧嘩したりと、喧嘩の程度や相手はさまざまでした。
喧嘩のあとの行動
ある日、喧嘩のあとでセキセイインコがつがい相手のところへすぐにとんでいくことに気がつきました。喧嘩のあとではつがい相手とどんな行動をしているのだろう。もしかしてなかなおりやなぐさめといった行動が見られるかもしれない。
そこで、セキセイインコの様子をしばらく録画して観察してみることに。録画した映像を見ながら「いつ」「どの個体が」「どの個体に対して」「どのような行動をしていたか」を時系列に沿って書き出してみることにしました。
時系列に書き出してみただけでは何が起こっているのかはまだ分かりません。「喧嘩のあとになかなおりやなぐさめがおこるのか」ということを調べるには、書き起こしたデータの集計や統計を通して仮説を検証する作業が必要です。
ところで、なかなおりやなぐさめで見られるやりとりを親和(しんわ)交渉と言います。ヒトなら握手やハグなど、トリなら羽づくろいや、嘴を近づけるというやりとりが親和交渉です。
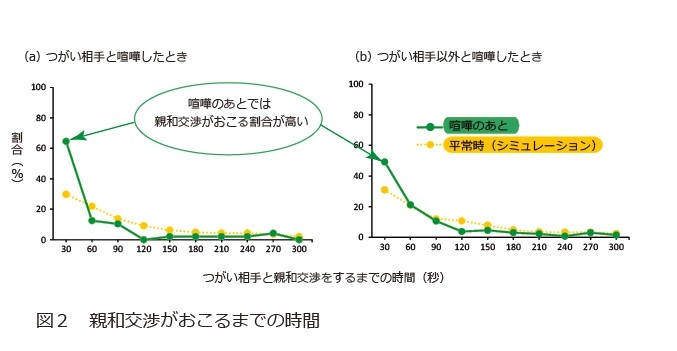
喧嘩のあとでは親和交渉が起こりやすい。つまり、喧嘩のあとでは親和交渉が起こるまでの時間が短くなるとの予測をもとに、実際のデータをつかって喧嘩から親和交渉までの時間を測定していきました。そして、シミュレーションによって(喧嘩が起きていない)平常時と比較しました。
その結果、喧嘩のあとでは親和交渉がおこるまでの時間が短くなっていたことが分かりました(図2)。
具体的に言えば、
(1)つがい相手と喧嘩するとそのあとつがい相手となかなおりをする
(2)つがい相手以外と喧嘩するとそのあとつがい相手と親和交渉をする
ことが分かりました(ちなみに(2)の一部のやりとりがなぐさめに該当します)。
ただし、すべてのつがいがこのような行動をしたわけではありませんでした。その理由はまだよく分かっていませんが、もしかしたらつがいの「絆」の違いによって、喧嘩のあとのフォローの仕方も違ってくるのかも?
カラスでの報告
調べてみると、カラス科のトリでも同じような報告があることが分かりました。ミヤマガラスは、つがい相手以外と喧嘩したあとつがい相手と親和交渉を行っていました [5]。コクマルガラスでも同じような報告がありました [6]。でもカラス科のトリのつがいでは、なかなおりは報告されていないのです。セキセイインコのつがいとは違って、カラス科のトリのつがいではめったに喧嘩がおこらないそうです。このことが、なかなおりがみられない理由のひとつ、と考えられています。
つまり、セキセイインコもカラスも喧嘩のあとに同じようなやりとりをします。でもその方法はちょっとずつ違うのです。
実験室での研究のすすめ
さて、私はこれまで実験室で飼育下のセキセイインコを対象に研究を行ってきました。「トリの研究をしています」と言うと、野外に出て野鳥を追いかけるイメージを持たれることが多くあります。
野外で鳥類の研究をしている研究者はたくさんいます。実験室で鳥類の研究をしている研究者もたくさんいます。野外と実験室の両方で研究をしている研究者もたくさんいます。実験室の研究の利点とはなんでしょうか。例えば、個体数、個体の年齢、観察時間など、野外ではコントロールできない様々な条件をコントロールして観察・実験できるということがあります(そして楽しい)。
大学や大学院に進学して鳥類の研究をしたいと思っている人がいたら、ぜひ実験室での研究も視野に入れて研究テーマを探してみてください。実験室ならではの、面白いテーマが見つかるかも!
【引用文献】
[1] Ikkatai, Y., Watanabe, S., & Izawa, E. I. (2016). Reconciliation and third-party affiliation in pair-bond budgerigars (Melopsittacus undulatus). 153, 1173 – 1193.
[2] Gallup, A. C., Swartwood, L., Militello, J., & Sackett, S. (2015). Experimental evidence of contagious yawning in budgerigars (Melopsittacus undulatus). Animal cognition, 18(5), 1051-1058.
[3] Farabaugh, S. M., Linzenbold, A., & Dooling, R. J. (1994). Vocal plasticity in budgerigars (Melopsittacus undulatus): evidence for social factors in the learning of contact calls. Journal of Comparative Psychology, 108(1), 81-92.
[4] Hile, A. G., Plummer, T. K., & Striedter, G. F. (2000). Male vocal imitation produces call convergence during pair bonding in budgerigars, Melopsittacus undulatus. Animal Behaviour, 59(6), 1209-1218.
[5] Seed, A. M., Clayton, N. S., & Emery, N. J. (2007). Postconflict third-party affiliation in rooks, Corvus frugilegus. Current Biology, 17(2), 152-158.
[6] Logan, C. J., Emery, N. J., & Clayton, N. S. (2013). Alternative behavioral measures of postconflict affiliation. Behavioral Ecology, 24(1),98-112.